情報・システム研究機構
データサイエンス共同利用基盤施設/国立遺伝学研究所
荒木 弘之
 小川智子先生が急逝されてもう4ヶ月がたつ。突然の訃報には驚いた。初めてお会いしてから40有余年、色々なことが思い起こされる。先生の学問的業績については篠原氏に譲り、私は人となりを偲ぶことにしたい。私は(多くの身近な人も)小川先生ではなく、智子さんと呼んでいたので、ここでもそれで通させていただく。
小川智子先生が急逝されてもう4ヶ月がたつ。突然の訃報には驚いた。初めてお会いしてから40有余年、色々なことが思い起こされる。先生の学問的業績については篠原氏に譲り、私は人となりを偲ぶことにしたい。私は(多くの身近な人も)小川先生ではなく、智子さんと呼んでいたので、ここでもそれで通させていただく。
私が大学3年時の遺伝学の実習は、バクテリオファージや大腸菌を用いたものであった。ファージの濃度を測定するため、ファージを含む液をガラスピペットを使って複数回希釈するのだが、そのデモンストレーションに登場されたのが智子さんである。ピペットの目盛を見ているのか分からないぐらい素早い。本人曰く、感覚でほぼ欲しい量吸い取れている。半信半疑ながら、すごい!と思った。研究室に入ると、大学院入試までの半年間は、実験よりお勉強をということで、机をあてがわれ勉強をしていた。その場所が智子さんの実験室兼居室であった。ある時、論文を読む演習の題材に智子さんの富澤研時代の論文(ラムダファージのDNA複製を調べたものでJournal of Molecular Biologyに出版された)を読んでいたら、後ろからちらっと見て、それね、3ヶ月間ぐらいのデータよと!なんかすごい。
recAタンパク質の精製を始められた時も、驚かされた。カラムクロマトグラフィー後の画分をアクリルアミド電気泳動で調べ、該当タンパク質があると思われるところを次のカラムにかけるという手順で進められていた。電気泳動の後の、染色・脱染の時間が長いと、電気泳動後、染色・脱染を高温でやり始めた。ようは煮るのである。でも、このおかげで急激に速くなり、複数のカラムを短時間で終えられることになった。ともかく頭が柔らかい!アイデアもそうなのです。
遺伝研では私が着任して、智子さんが退官されるまでの3年間お世話になった。ラボの運営の仕方などなどです。ジャーナルクラブも仕事のセミナーも3年間一緒にやらせていただいた。いつも固定観念にとらわれることなく見ておられたことがよくわかった。Mre11がRad50, Nbs1と複合体を作っていることが分からなかった時に、Mre11のDNA結合能や2本鎖同士を近くに連れてくることなどに大変興味を持って研究をされていた。当時、何かのアーティファクトかもと思っていた我が身が恥ずかしい限りである。私のところでは、染色体DNA複製の開始と細胞周期チェックポイントに関与する出芽酵母のDpb11(哺乳類ではTopBP1)を解析していた。勿論細胞には必須だが、最初に分離した温度感受性変異は764アミノ酸のうちC末の182アミノ酸を欠くものである。そして、この変異は許容温度でも弱い細胞周期チェックポイントの異常を示した。そのため、Dpb11のチェックポイントの機能領域を探そうとしていた。C末欠損で温度感受性になるので、この欠損がN末側に影響を与えると考え調べていたのだが、智子さん曰く、C末の欠損したところをどうして調べないの。でも、表現型が弱くて・・・。結局わかってみるとやっぱりそうで、そこにはチェックポイントキナーゼMec1(哺乳類のATR)を活性化する配列があることを他のグループが見つけた。何とも残念なのだが、智子さんに指摘されていたことは今でもよく覚えている。言い訳をさせていただければ、並列に働く系があって、それを除かないと解析が難しかったのです。的確な指摘でした。
研究はやれることをとことんやる。もう、これ以上考えてやることがないというギリギリのところまでやらないと優れた仕事にはならないと常々言っておられた。その通りだと思う。最終講義の最後に、若い人にと言われたことが心に残っている。当時のスライドには「直接的証明のできる実験/目的を明確に/その時点でできることは行う/他人の実験をよく見る/人の意見を聞く/良く考え、実行すること/多角的データ解析/指導者を選ぶ」とあります。
退官される年に日本分子生物学会の会長職につかれた。その際、庶務幹事を任せられ、二人で行動をともにすることも多かった。在任期間には、三菱化学奨励賞の設置、学協会の男女共同参画の立ち上げ等を行なった。一緒に相談したこともあったし、一任されて動いたこともあったが、智子さん本来のはっきり物を言う性格のため、進めやすかった。
まとまりのない文になりましたが、小川智子先生のご冥福を祈ります。先生の残されたものの一部でも、我々が引き継ぐことが重要であろうかと思います。
大阪大学 蛋白質研究所
篠原 彰
小川智子先生(国立遺伝学研究所教授、分子生物学会第12期会長)が2020年12月24日金曜日の夕刻に、先生の地元、盛岡で83歳という年齢で永眠されました。コロナ禍ということもあり、親族のみでの葬儀(告別式は12月28日)となり、そのお顔を拝見することなく、永遠の別れとなってしまいました。喪失感も、全く現実感がともなわない。2021年4月この原稿を書いている今でも、智子先生が傍で「篠原くん、何しているの?しっかりしなさい」と声が聞こえてきそうな気がしてならないのです。この場を借りて、智子先生に感謝の意を表したく、加えて智子先生の研究業績、人柄を皆さんにお伝えできればと思います。
小川智子先生を一言で表すなら、すごい、これに尽きる。多くの研究者の方々は勿論、智子先生が素晴らしい研究者だと認識されていると思います。それでも、智子先生の研究者としての凄さが過小評価されているからこそ、その凄さを伝えるのがこの文を書く僕の役目だと思っています。智子先生は1960年に共立薬科大学(現在の慶應大学薬学部)を卒業され、卒業後、国立予防衛生研究所(予研―現在の感染研)に就職されました。その後、1968年に東京大学で学位を取得しています。学歴で人を判断すべきではないですが、日本が戦後の復興を遂げている中で、大学院での教育、研究の経験が全くないまま、一流の研究者まで上り詰めた女性という意味では稀有ではないか、と思います。
智子先生は予研では富澤純一博士(元国立遺伝研所長、初代Genes-to-Cells編集長、ColE1プラスミドの複製制御に関わる制御性の小分子RNAの発見など)のもとで、研究を始められました。当時、富澤先生は大腸菌やファージを用いた分子遺伝学(分子生物学の原型)を積極的に研究に取り入れたDNA複製や組換えに関する研究を行い、世界的な業績を上げていました。智子先生は大腸菌のウイルスであるラムダファージのDNA複製中間体を世界に先駆け電子顕微鏡で観察し (Ogawa T, et al. PNAS, 60, 861-85, 1968)、その成果をもとに東京大学で学位を取得されました。その後、富澤先生が大阪大学理学部生物学科に異動するのに伴い、小川英行先生と一緒に、大阪大学に助手として赴任されました。学園紛争の中、1970年から2年間は米国のマサチューセッツ工科大学に研究留学されました。富澤先生が1971年に米国NIHに異動された後に帰国されて、1975年に小川英行先生が教授になられ講座を主宰される中で、智子先生は1984年に講師となられました。その後、1995年に国立遺伝研の教授として異動され、それから大阪大学の教授も兼任されました。智子先生は大腸菌研究の分野ではもともと著名でしたが、この時を前後して、より多くの方々に小川智子先生の名前が知られるようになったと記憶しています。
私、篠原は1985年に学部学生として研究室に配属され、智子先生を指導教官として研究を開始しました。その当時の、小川英行先生(現 大阪大学名誉教授; 岩手看護専門学校校長)、智子先生と伊藤建夫先生(現信州大学教授 名誉教授)、堀井俊宏先生(現 大阪大学特任教授)がスタッフだった小川研は大腸菌の遺伝学、生化学、出芽酵母の分子遺伝学、分子生物学を融合した多彩な研究論を用いて、DNA複製や組換えの分子メカニズムを研究していました。ですので、実験セミナーで出てくる研究の話題は多岐に渡って、刺激の多いものであったことは言うに及びません。学生達はその当時、智子先生のことを、親しみを込めて、“智子リン”と呼んでいました。ことあるごとに、智子リンはすごい、それが我々学生の口癖でした。
智子先生は大阪大学に移って来てからは、大腸菌のrecA遺伝子、RecAタンパク質の機能解析を中心に研究を展開されました。当時、遺伝子組換え技術、例えば、遺伝子クローニングやDNA塩基配列の決定方法は世に出て間もない頃でしたが積極的に取り入れられていました。研究室で制限酵素BamHIやT4 ligaseを精製して、形質導入を用いてrecA遺伝子を含むラムダファージから得て、そのファージからrecA遺伝子を含むDNA断片を、ColE1プラスミド上へクローニングを行い、電子顕微鏡を使って、Rループ法で遺伝子の位置を特定したわけですから驚きです。大腸菌でDNA損傷が誘導されることで誘発される38kDのタンパク質(プロテインXとその当時は言われていた)がrecA遺伝子の産物であることを示したのです。さらに、いわゆる今の組換えタンパク質の量産系がない状態で、RecAタンパク質を精製して、オリゴマー形成活性があり、1本鎖DNA依存性のATP加水分解(ATPase)活性があることを証明しました(Ogawa T et .al. Cold Spring Harbor Symp Quant Bio, 1979)。無いなら習う、取る、本当にありとあらゆる手立てを使って、自分の知りたいこと、新しいことを追求して、明らかにする、素晴らしい一連の仕事だと思います。あの当時に分野を超えた統合生物学を先駆的に実施してたことは驚きです。私が学生だった1980年の後半には、先述のrecA遺伝子の研究の進展についての話と、1979年のCold Spring Harbor Symposiumで講演をして賞賛を浴びたことを誇らしげに話してくれたことが今でも思い出されます。多分、そこには我々の知らない大変な苦労があったのでしょうが、厳しい中でも研究を楽しんでいる智子先生の姿が今でも残っています。RecAタンパク質の活性同定の後、recA遺伝子とその発現のリプレッサーであるLexA遺伝子(酵母two-hybrid法でDNA結合ドメインとして使われている)のDNA塩基配列を決定して(Horii et al, PNAS USA, 1980; Horii et al. Cell, 1981a)、一本鎖DNA に結合したRecAがLexAタンパク質を切断する活性(厳密にはLexAの自己切断活性を活性化する)を見つけ、大腸菌のDNA損傷応答をin vitroで再構成することで、DNA損傷応答の分子メカニズムを見事に明らかにしました(Horii et al. Cell, 1981b)。
新しいことにトライすることも好きなのが智子先生なのです。学部の3年生の実習でrecA変異のサプレッサーを単離して、放射性同位体を使ったサンガー法でそのDNA塩基配列を決めるなんていう実験を1984年に実施しているのです。RecAの機能を知る為にはその構造を知りたい、そのためにタンパク質の機能をお互いに相補する2つのアミノ酸変化の相関から、RecAの構造が分からないか、と考えて、遺伝子内のサプレッサーから分子遺伝学でタンパク質の構造を知ろうという壮大な計画を学部の学生にやらせるのですから。無茶過ぎです。
RecAタンパク質のATPase活性の発見が大きな契機となり、大腸菌における相同組換えの研究が爆発的に進みました。これらrecA遺伝子の研究に加えて、智子先生の大きな業績として、真核生物で初めてRecAと相同なRad51を見つけ、酵母からヒトまで普遍的に存在する遺伝子であることを見つけたことが挙げられます。それが大きな契機になり、真核生物の相同組換え機構や、遺伝子ターゲッティングやゲノム編集といった、組換えに関わる技術開発にも大きな貢献をしました。1980年代中頃から後半の時期には、ヒトなどの高等真核生物ではRecAのようなスーパーな組換え酵素はないと言う考え方が蔓延していました。高等真核生物では細菌や酵母に比べて、相同組換え(ジーンターゲッティング)の効率が悪いこと、生化学的にRecAと同じように試験管内でDNA鎖交換活性を持つタンパク質の同定が国内外で多くの研究者によって報告されましたが、ヒト細胞は言うに及ばす、組換え活性の高い酵母からですら、精製されてくるタンパク質はすべてATP非依存性のDNA鎖交換活性を示すタンパク質で、その配列もRecAとは異なるものでした。さらに決定的だったのは小川研でパン酵母の相同組換えに最も大切だと考えられてきたRAD52遺伝子をクローニングしていたのですが、Rad52のアミノ酸配列がRecAと全く異なっていて、ATPに依存した活性は同定できていませんでした。そのような状況で、酵母を含め、真核生物にはRecA様のタンパク質は存在しないのでは?という状況だったと言えます。ただ、一方で小川研では「ATPに依存する活性を持つのがRecAである」という見方が当たり前のようにありました。智子先生もそう信じていたはずです。ですから、智子先生の下で教育を受けた私が真核生物でRecAと同じATP依存性の組換え酵素を見つけたいと思って研究を始めたのは自然の成り行きだったかもしれません。智子先生は常に正しい筋道、行先を与えてくれました。修士時代のrecAの研究から新しい研究テーマをしたいと言い出し、智子先生と激論しました。その時、過去の成果や研究に対する私の不遜な態度を厳しい言葉で指導されました。そのような議論の後、新しい方向性をたどる中で、たまたまRAD51遺伝子に出会い、そのDNA塩基配列を決め、Rad51タンパク質のアミノ酸配列を見出しました。その当時はBLASTなどの検索サイトは存在せずに、会社が販売しているデータベース検索ソフトを用いました。その時に検索結果として出てきたトップが大腸菌の転写終結因子(ロー因子)と複製開始タンパク質のDnaBでした(これらは2つともATP加水分解依存性のヘリカーゼ)。RecAはというと、トップ30にも入っていませんでした。ただ、Rad51は配列から見て、間違いなくATP結合モチーフ(WalkerのA型、B型)を有していました。つまり、ATP結合、加水分解活性を持つのは間違いない、と。そこで、役に立ったのがrecAのサプレッサー変異による遺伝的タンパク質の構造解析でした。私は博士の1年まではその大腸菌プロジェクトに関わり、智子先生と一緒に、穴が開くほどRecAの配列を見ていました。気がついていたのはWalkerのB型配列(xxxxD;xは疎水性アミノ酸)の特徴でした。智子先生はこのモチーフを当初からなぜか指摘していました。この配列はタンパク質のサブファミリー毎に特徴的であることが知られていました。有名なタンパク質としてxxxxDEAD boxを持つヘリカーゼが上げられます。RecA はxxxxD“S”で、他のATP結合タンパク質とは似ていない配列を有していたことを覚えていました。そして、Rad51のWalkerのB型配列はRecAのxxxxD“S”型で同じ。もしやと思って、2つのアミノ酸配列を目で並べると、意外と並ぶのです(相同性は20%も満たない、ギャップも多く存在し、非常に弱いものでした)。Rad51が酵母のRecA相当タンパク質では?と思った瞬間でした。Rad51の配列がRecAと似ていると、智子先生に話した時はすごく喜んでおられました。
その後、Rad51の解析は進んでいったのですが、たまたま、Harvard大学のNancy Kleckner博士の研究室でも酵母のRecA相同遺伝子を見つけたと言う話が流れてきました。その当時(今もですが)Kleckner博士は組換え分野の新進気鋭の研究者で、トランスポゾンTn10や減数分裂期組換えで素晴らしい仕事を連発していました。その情報を聞いた瞬間は敗北という言葉が浮かんだのは言うまでもありません。ただ、その時に智子先生のすごさが発揮されたのです。Kleckner博士は智子先生の知り合いだったのです。しかも、「Nancyは私が研究者に導いた」と智子先生は豪語していました。智子先生がMITに留学していた時に、Kleckner博士は同じ研究室に在籍していて、智子先生から分子遺伝学のイロハを教えてあげたのだと。智子先生に言わせるとNancyは私が助けたとなるのですが、実際に、Nancyの智子先生への対応を見ていると、本当なのだと確信しています。ですので、Nancy(智子先生との絆)を信じて、こちらが持っているRad51のアミノ酸配列をHarvard大学にFAXしたのです。もし同じだったらと、大きな不安を抱えて。翌日の、FAXの返信を見てみると、我々が見つけたRad51と、NancyとDoug Bishop博士が見つけた遺伝子(Dmc1)は違っている、と言うことが分かりました。さらに驚きだったのはRad51とDmc1は双子、似ているけど違うのです(相同性は約50%)。つまり、酵母は2つRecA相同遺伝子を持っている、体細胞分裂期型と減数分裂期型、と大きな発見に繋がったのです。それを契機に2つの論文が一緒に発表されました。智子先生のKleckner博士の繋がり、絆がなかったら、そして、智子先生がRecAのATP加水分解活性を見つけていなかったら、起こり得なかった研究展開だと思っています。
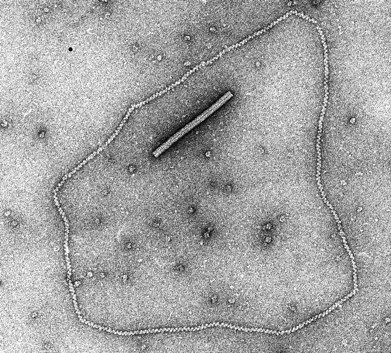
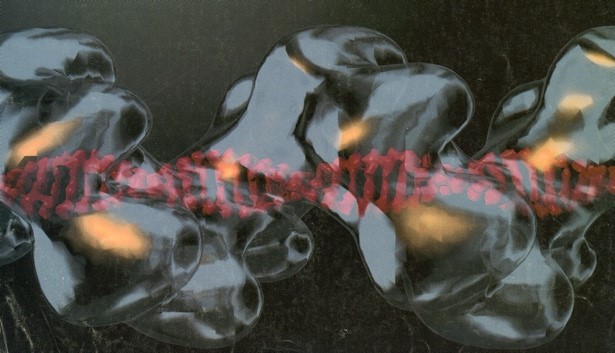
智子先生がすごいのは徹底的な現場主義だと言う点です。大阪大学にいた当時から実験が大好きな方で、かつ実験が上手い(観察眼が鋭いし、賢いし、早いし、手際がいい)のです。智子先生の頭の回転の速さは、智子先生が父と仰いでいた富澤先生譲りではないでしょうか。何より、実験を沢山するのです、私が学生だった当時、智子先生はもう五十歳に近い年齢でしたが、深夜まで実験していました。時には午前様になる我々学生よりも遅くまで、朝帰りなんってこともあったと記憶しています。夢中になると止まらない、好きなのですね、実験が本当に。その腕が発揮されたのは勿論、電子顕微鏡によるRad51-DNA複合体の構造解析でした。Rad51の研究の新しい展開として、Rad51の活性や構造を決めるという方針でタンパク質を精製していました。阪大でも電子顕微鏡でRad51-DNA複合体を見ようと私自身もトライしましたが、カーボンの薄膜から作る職人芸に驚き、かつ、実際に見える緑(電子線の像)の砂の嵐の中でのものを見ることはほぼ不可能だと思っていました。そんな中で智子先生が国際会議で知り合った、電子顕微鏡の専門家であったミネソタ大学のEd Egelman博士と共同研究の話が持ち上がりました。先の研究で電子顕微鏡にトラウマがあった僕ではなく、智子先生が、私が酵母から精製したてのRad51を持ち、ミネソタに行くことになりました。向こうに行って、1ヶ月もしないうちに、「篠原くん、Rad51はRecAと同じ、Rad51、区別がつかない」、という連絡が来ました。そして、発表したのがRad51-DNA複合体の構造でした(図; Ogawa T., et al. Science, 259, 2856-, 1993)。その美しさ、そして何よりRecA-DNA複合体と同じであることに驚愕したと同時に、俺が行けば、と思ったものでした。でも、智子先生の腕と経験があったからこそできた仕事なのだとその当時は驕りもあって思えなかったのでしょう。
|
|
左―Rad51-DNAの電子顕微鏡写真;
右―再構成したRad51-DNAフィラメントの構造(DNAが1.8倍伸張している);
Ogawa T., et al. Science, 259, 2856-, 1993
智子先生はその不思議な魅力から、いろいろな人との繋がりがありました。その中で、上述したNancy Kleckner博士、MITでKleckner博士と同期だったIra Herskowitz博士(出芽酵母の分子遺伝学の大家)とも仲がいいし、Herskowitz博士とUCSFで同僚だったBruce Alberts博士(あの有名な教科書-The Cell/細胞の分子生物学の最初の著者の一人)とも仲間です。Alberts博士は大腸菌のファージ、T4ファージのDNA複製、組換えの専門家です。特に今では当たり前ですが、タンパク質―タンパク質の相互作用の重要性という視点から複製や組換えの研究をやっていました。学生だった僕を外国に連れ出し(イメージアナライザーの購入のための説明を受けるためにアメリカに学生を行かせるという無茶振り)、その際にAlberts博士のラボでT4ファージの複製や組換えに関わるタンパク質群の、複雑、かつ精巧なタンパク質―タンパク質の相互作用の解析について話をして、それが生かされたのがRad51の論文です。あの論文ではRad51がRecA相同遺伝子であることが強調されますが、あの論文ではRad51がRad52に結合するということも示しています。それを試そうと思ったのはAlberts博士から、タンパク質―タンパク質の相互作用のことを聞いていたからです。つまり、智子先生はすでにタンパク質―タンパク質の相互作用の重要性に気がついていたのでしょう。智子先生がいなかったら生まれなかった研究展開だと言えます。この酵母のRad51の遺伝子の配列、たかが4,500塩基対ですが、その当時のDNA配列の決定となると大騒ぎです。そこでも、智子先生はその当時出始めたばかりの蛍光色素を用いたDNAシークエンサー(当時のABI社製)を使うことを提案したのです。しかも、デモと称してサンプルをRad51のDNA断片も含めて沢山解析して貰いました。そのおかげで、その当時としては効率的に配列を決定できたと思います。すごい情報網です。そして新しい物好き。
 その後、酵母のRad51からヒトのRAD51遺伝子をクローニングするのですが、その時も一波乱がおきました。まずクローニングする際には遺伝子が乗ったラムダファージを大量に増やす必要があります。僕はなぜかラムダファージを増やすのが上手くなく、智子先生が毎日のように50cmに近いガラスのシャーレに生やした大腸菌からラムダファージを集めてくれました。一ヶ月近くも一緒に夜遅くまで実験していました。あの時の先生の協力がなかったら、RAD51の配列は順調には決まらなかったでしょう。ヒトのRAD51遺伝子のクローニングの計画は極秘に進めていたのですが、なぜか情報が漏れて、国内の他のグループも見つけ、論文にするという話が持ち上がりました。その時、智子先生と二人で、その研究室と話し合いに行った時、喧嘩を売ろうとする僕をなだめつつ、サポートしてくれた姿は今でも大切な思い出になっています。その研究を論文にする時もどのような雑誌に投稿したらよいかなど、智子先生がHerskowitz博士と話をつけたりして、なんとか発表出来ました(Shinohara et al. Nature Genetics, 1993年)。智子先生の人脈と人柄がなかったら、うまく発表できなかった仕事だと思っています。
その後、酵母のRad51からヒトのRAD51遺伝子をクローニングするのですが、その時も一波乱がおきました。まずクローニングする際には遺伝子が乗ったラムダファージを大量に増やす必要があります。僕はなぜかラムダファージを増やすのが上手くなく、智子先生が毎日のように50cmに近いガラスのシャーレに生やした大腸菌からラムダファージを集めてくれました。一ヶ月近くも一緒に夜遅くまで実験していました。あの時の先生の協力がなかったら、RAD51の配列は順調には決まらなかったでしょう。ヒトのRAD51遺伝子のクローニングの計画は極秘に進めていたのですが、なぜか情報が漏れて、国内の他のグループも見つけ、論文にするという話が持ち上がりました。その時、智子先生と二人で、その研究室と話し合いに行った時、喧嘩を売ろうとする僕をなだめつつ、サポートしてくれた姿は今でも大切な思い出になっています。その研究を論文にする時もどのような雑誌に投稿したらよいかなど、智子先生がHerskowitz博士と話をつけたりして、なんとか発表出来ました(Shinohara et al. Nature Genetics, 1993年)。智子先生の人脈と人柄がなかったら、うまく発表できなかった仕事だと思っています。
智子先生のことを語り出したら、キリがないのですが、もう1つ話をさせてください。RAD51の相同遺伝子をヒトでクローニングして、当然、普通の人はヒト細胞やマウスなどで解析となるのですが、智子先生は違うのです。ある日、突然(多分、突然ではないのでしょうが)、「篠原くん、ユリよ、ユリ」、と言い出したのです。はぁ、と呆れていると、次の日には和歌山のユリ農家まで足を運んで、研究室の智子先生の部屋は青いバケツにはいった蕾がついたユリで一杯になりました。で、ユリ(植物の中でも染色体が大きく細胞生物学には適していて、蕾の大きさで減数分裂期のステージが分けることが出来る)の雄蕊の減数分裂の染色体を見ようと思い付いたようです。ここで、すごいのはそればかりでなく、作りたてのヒトRAD51の抗体で蛍光染色をやると言い始めたのです。RAD51は酵母からヒトまで保存するという信念でクローニングした僕ですら、その発想はありません。ヒトの抗体がユリで働く、そんな発想はどこから生まれてくるのか。そして、驚異的なことに、ヒトのRAD51の抗体で減数分裂期の染色体上のユリRAD51が染まり、fociとして検出できたのです(Terasawa et al. Genes Dev. 9, 925-, 1995―表紙の写真)。その像を見せられた時は圧倒されました。 呆れつつも、この人はすごい、と。
その後、1995年に智子先生は国立遺伝研教授として赴任されました。富澤先生の近くに行きたいという思いがあったのだと思います。その当時の智子先生と同じ年齢になり、改めて今の自分と智子先生を比較すると、惨敗です。何も勝てていません。智子先生ごめんなさい、です。2002年に退職されて、ご実家のある盛岡に戻られて、その看護短期大学で、看護師の育成、教育に力を入れつつ、研究を続けられました。富澤先生のもとにはかなりの回数で毎年訪れられていました。本当に尊敬していたのだと思っています。
やはり、智子先生の最もすごいところは研究業績であることは言うまでもありません。智子先生は一見、豪快に見え、口下手だから誤解されがちですが、本当は繊細で、誰にでも優しい、人間的にも素晴らしい方です。理学部の生物学科のレクレーションのバレーボール大会に出て、アキレス腱を切った逸話。国際学会の懇親会のダンスパーティーで外国の方と社交ダンスを踊った姿。どんなに紙面を頂いても、書き足りるものではありません。何より、国際的にも、国内的にも過小評価されているということは残念でなりません。弟子である僕がその後、智子先生を押し上げることをしなかった、それが大きな慚愧として残っています。ノーベル賞はDNA複製で2回、DNA修復で受賞者が出ています。組換えで受賞がないのは同じ分野に働くものとして不思議でなりません。仮に、組換えが受賞したら、Nancy Kleckner博士と智子先生がノーベル賞を貰ってもおかしくないと、ずっと昨年までは思っていました。でも、そう言う夢を見ることは残念ながら無くなりました。それでもこの文章を読む方々には、改めて、小川智子先生という稀有の科学者がいたことを知って頂きたいと思います。
智子先生、ありがとうございました。そして、お疲れ様でした。
Ogawa, T., Tomizawa, J.I., and Fuke, M. (1968). Replication of bacteriophage DNA, II. Structure of replicating DNA of phage lambda. Proc Natl Acad Sci U S A 60, 861-865.
Ogawa, T., Wabiko, H., Tsurimoto, T., Horii, T., Masukata, H., and Ogawa, H. (1979). Characteristics of purified recA protein and the regulation of its synthesis in vivo. Cold Spring Harb Symp Quant Biol 43 Pt 2, 909-915.
Shinohara, A., Ogawa H., and Ogawa T. (1992). Rad51 protein involved in repair and recombination in S. cerevisiae is a RecA-like protein. Cell 69, 457-470.
Ogawa, T., Yu, X., Shinohara, A., and Egelman, E.H. (1993). Similarity of the yeast RAD51 filament to the bacterial RecA filament. Science 259, 1896-1899.
Terasawa, M., Shinohara, A., Hotta, Y., Ogawa H., and Ogawa T. (1995). Localization of RecA-like recombination proteins on chromosomes of the lily at various stage of meiotic stages. Genes & Dev. 9, 925-934.
